I. Типы межвидовых отношений. Конкуренция.
Типы взаимодействия между двумя видами
Живые организмы не могут существовать сами по себе. Они связаны воедино разнообразными отношениями, вся полнота которых выявляется лишь при анализе экосистемы как целого. Живые существа зависимы от своего окружения, поскольку им надо питаться, расселяться, защищаться от хищников и т. д. Виды по-разному влияют друг на друга: конкурируют с соседями за пищу и выделяют ядовитые вещества, оказываются полезными друг для друга или «эксплуатируют» другие виды. Сущность межвидовой конкуренции заключается в том, что у особей одного вида уменьшается плодовитость, выживаемость или скорость роста в результате использования ресурса или интерференции со стороны особей другого вида. Однако за этой простой формулировкой кроется большое количество самых разнообразных нюансов. Влияние межвидовой конкуренции на динамику численности популяции конкурирующих видов многолико. Динамика в свою очередь может оказывать влияние на распределение видов и их эволюцию.
Конкуренция возникает тогда, когда два или большее число организмов получают ресурс из источника, который для всех явно недостаточен. Иными словами, конкуренция возникает только в том случае, если ресурс ограничен. Если же ресурс имеется в избытке, то два вида даже с очень сходными потребностями конкурировать не будут. Это может наблюдаться, например, при регулировании численности видов хищниками или паразитами.
нейтрализм,
взаимное конкурентное подавление,
аменсализм,
Таблица 1 Анализ взаимодействий популяций двух видов 1
|
Тип взаимодействий 2 |
Виды |
Общий характер взаимодействия |
|
|
1 |
2 |
||
|
1. Нейтрализм |
0 |
0 |
Ни одна популяция не влияет на другую |
|
2. Конкуренция, непосредственное взаимодействие |
— |
— |
Прямое взаимное подавление обоих видов |
|
3. Конкуренция, взаимодейсвия из-за ресусрсов |
— |
— |
Непрямое подавеление при деффиците общего ресурса |
|
4. Аменсализм |
— |
0 |
Популяция 2 подавляет популяцию 1, но сама не испытывает отрицательного воздействия |
|
5. Паразитизм |
+ |
— |
Популяция паразита (1) состоит из меньших по величине особей, чем популяция хозяина |
|
6. Хищнечество (и поедание растений) |
+ |
— |
Особи хищников (1) обычно крупнее, чем особи жертвы (2) |
|
7. Комменсализм |
+ |
0 |
Популяция 1, комменсал, получает пользу от объедения; популяции 2 это объедение безразлично |
|
8. Протокооперация |
+ |
+ |
Взаимодействие благоприятно для обоих видов, но не обязательно |
|
9. Мутуализм |
+ |
+ |
Вазимодействие благоприятно для обоих видов и обязательно |
1. 0 означает отсутствие значимых взаимодействий; + означает улучшение роста, выживания и другие выгоды для популяции ( к уравнению роста прибавляется положительный член);
- означает замедление роста и ухудшение других характеристик (к уравнению роста добавляется отрицательный член).
2. Типы 2-4 можно считать «отрицательными взаимоотношениями», типы 7-9 – «положительными взаимоотношениями», а типы 5 и 6 можно относить к обеим этим группам.
Следует подчеркнуть три принципа, основанные на этих категориях:
1. Негативные взаимодействия проявляются на начальных стадиях развития сообщества или в нарушенных природных условиях, где высокая смертность нейтрализуется r-отбором.
2. В процессе эволюции и развития экосистем обнаруживается тенденция к уменьшению роли отрицательных взаимодействий за счет положительных, повышающих выживание взаимодействующих видов,
3. В недавно сформировавшихся или новых ассоциациях вероятность возникновения сильных отрицательных взаимодействий больше, чем в старых ассоциациях.
Одна популяция часто влияет на скорость роста или смертность другой. Так, члены одной популяции могут поедать членов другой популяции, конкурировать с ними за пищу, выделять вредные вещества или взаимодействовать с ними другим способом. Точно так же популяции могут быть полезными друг для друга, причем польза в одних случаях оказывается обоюдной, а в других — односторонней. Как было показано в таблице 1, взаимодействия такого рода подразделяются на несколько категорий.
Все эти популяционные взаимодействия можно изучать на примере некоего среднего сообщества. Тип взаимодействий конкретной пары видов может изменяться в различных условиях или в зависимости от стадии жизненных циклов. Так, в одно время у двух видов может быть выявлен паразитизм, в другое время — комменсализм, а в какой-то третий период жизни они могут быть полностью нейтральны друг к другу. Упрощенные сообщества и лабораторные эксперименты позволяют экологам получать всего один из многих типов взаимодействий и изучать его количественно. Дедуктивные математические модели, построенные на основе таких исследований, дают возможность анализировать факторы, которые обычно трудно отдифференцировать один от другого.
Для выяснения действий различных факторов в сложных природных ситуациях, а также для более точного определения понятий и большей четкости рассуждении полезно пользоваться «моделями» в форме уравнений. Если рост одной популяции можно описать с помощью уравнения, то влияние другой популяции можно выразить членом, который изменяет рост первой популяции. В зависимости от типа взаимодействия в уравнение можно подставлять разные члены. Например, при наличии конкуренции скорость роста каждой популяции равна скорости нелимитируемого роста минус влияние собственной численности (которое растет с ростом численности популяции) и минус величина, характеризующая отрицательное влияние другого вида, N 2 (которая также растет по мере роста численностей обоих видов N1 и N2 ), или
|
dN 1 |
= |
rN 1 |
— |
r |
N 1 2 |
— |
CN 2 N1 |
|
|
dt |
K |
|||||||
|
Скорость роста |
= |
Нелимитиру- емый рост |
— |
Влияние собственной численности |
— |
Отрицательное влияние другого вида |
||
Когда виды двух взаимодействующих популяций оказывают друг на друга благоприятное, а не повреждающее влияние, в уравнение вводят положительный член. В таких случаях обе популяции растут и процветают, достигая равновесных уровней, что благоприятно для обоих видов. Если для роста и выживания каждой из популяций необходимо их взаимное влияние друг на друга, то такие взаимоотношения называются мутуализмом. Если же, с другой стороны, эти благоприятные влияния лишь вызывают увеличение размеров популяции или скорости ее роста, но не являются необходимыми для ее роста и выживания, то такое взаимодействие соответствует кооперации или протокооперации. (Поскольку такая кооперация не есть следствие сознательной или «разумной» деятельности, предпочтительнее пользоваться последним термином.) И мутуализм, и протокооперация приводят к сходному результату: рост популяции в отсутствие другой либо замедлен, либо равен нулю. По достижения равновесия обе популяции в продолжают сосуществовать, обычно сохраняя определенной соотношение.
Рассмотрение популяционных взаимодействий так, как это сделано в таблице 1 или на основе уравнений роста, позволяет избежать противоречий, нередко возникающих при обсуждении терминов или определений как таковых. Например, термин «симбиоз» иногда употребляют в том же смысле, что и мутуализм, а иногда его понимают более широко, включая в него также комменсализм и паразитизм. Поскольку слово симбиоз буквально означает «Жить вместе», его следует применять в широком смысле, безотносительно к природе взаимосвязи. Термин «паразит» многие используют применительно к любому мелкому организму, живущему на поверхности или внутри другого организма, независимо от того, положительно, отрицательно или безразлично его влияние. В таком же смысле трактуется и предмет науки паразитология. Для одного и того же типа взаимодействий предлагались разные термины, что только увеличивало путаницу. Однако, когда отношения представлены в виде схемы, обычно не остается сомнений, к какому типу относить рассматриваемое взаимодействие; слово или «ярлык» оказывается в таком случае вторичным относительно механизма и его результата.
Следует обратить внимание на то, что слово «вредоносный» не использовалось в описании отрицательных взаимодействий. Конкуренция и хищничество снижают скорость роста популяций, испытывающих эти влияния, но этот эффект не обязательно вреден, если рассматривать его с точки зрения выживания популяции на протяжении длительного времени или с точки зрения ее эволюции. Отрицательные взаимодействия фактически могут ускорять естественный отбор, приводя к возникновению новых адаптации. Хищники и паразиты часто полезны для популяций, не имеющих механизмов саморегуляции для предотвращения перенаселения, следствием которого могло бы быть самоуничтожение таких популяций.
принцип конкурентного исключения.
принцип Гаузе
Tribolium
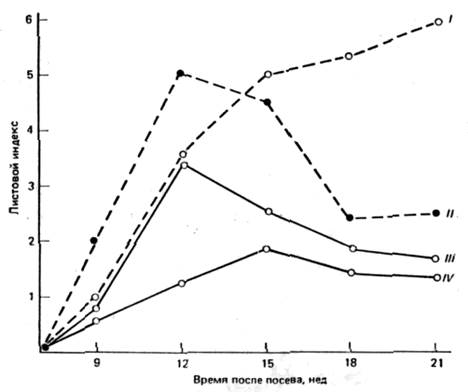
Рис. 1. Сосуществование видов клевера ( Trifolium ). На графике показан рост популяций двух видов в чистых (т, е. представленных только одним видом) и смешанных травостоях. Следует отметить, что внимание на то, что кривые роста двух видов в чистых травостоях различны и в разное время достигают зрелости. Это и другие различия позволяют видам сосуществовать в смешанных травостоях, но при этом их плотность понижена, так как они в одинаковой степени мешают друг другу. Листовой индекс, который используется для оценки плотности биомассы, представляет собой отношение площади поверхности листьев к поверхности почвы (см2 / см2 ).
I – T . fragiferum , чистый травостой; II — Т. repens , чистый травостой; III — T . repens , смешанный травостой; IV— T . fragiferum , смешанный травостой.
Tribolium
Таблица 2.
|
«Климат» |
Температура, о С |
Относительная влажность, % |
Исход межвидовой конкуренции 1 , % |
|
|
ПобеждаетТ.castaneum |
Побеждает Т.confusum |
|||
|
Жаркий влажный |
34 |
70 |
100 |
0 |
|
Жаркий сухой |
34 |
30 |
10 |
90 |
|
Теплый влажный |
29 |
70 |
86 |
14 |
|
Теплый сухой |
29 |
30 |
13 |
87 |
|
Холодный влажный |
24 |
70 |
31 |
69 |
|
Холодный сухой |
24 |
30 |
0 |
100 |
1. Каждый из 6 вариантов опытов проведен в 20—30 повторностях. В чистой -культуре каждый вид выживает при любых сочетаниях условий, однако при совместном культивировании двух видов выживает только один. В процентах выражено относительное число повторностей, в которых сохранялся только один вид, а другой элиминировался.
Tribolium
Tribolium
Trifolium
При изучении литературы, посвященной конкуренции, складывается общее впечатление, что в системах, где отсутствуют или понижены иммиграция и эмиграция, происходит более жестокая конкуренция и более вероятно конкурентное исключение. К числу таких систем следует отнести лабораторные культуры, острова или другие природные ситуации с трудно преодолимыми преградами для входа и выхода. В обычных природных открытых системах вероятность сосуществования выше.
Tribolium
то выживут обе популяции. Таким образом, если в среде имеются убежища, позволяющие скрываться от прямого воздействия (в данном случае хищничества), то конкуренция снижается настолько, что сохраняются оба вида.
Но довольно лабораторных примеров. Совершенно очевидно, что скученность в лабораторных экспериментах может быть весьма значительной, что приводит к чрезмерной конкуренции. В полевых исследованиях межвидовая конкуренция была подробно изучена у растений; результаты этих исследований позволили прийти к выводу (в настоящее время этот вывод является общепринятым) , что конкуренция — важный фактор, вызывающий смену видов. Кивер (Keever, 1955) описал случай, когда залежь первого года была почти целиком занята чистым травостоем высокого сорняка, в дальнейшем его постепенно вытеснил отсюда другой вид, ранее не известный в этих местах. Эти два вида, хотя и принадлежат к разным родам, имеют очень сходные жизненные циклы (время цветения и созревания семян) и Жизненные формы, попадали в условия интенсивной конкуренции. Тщательные последующие исследования этих залежных полей показали, что новый пришелец не вытеснял растущий здесь ранее вид; оказалось, что оба вида сосуществуют, но их численное соотношение зависит от почвы, времени и степени нарушений.
Следующий пример относится к двум видам сухопутных саламандр Plethodon glutinosus и P. jоrdani, обитающих в южной части Аппалачских гор в США. Обычно P. jоrdani встречается на больших высотах, чем P. glutinosus, но в некоторых районах зоны их обитания перекрываются . Хейрстон (Hairston, 1980) провел эксперимент на двух участках, один их которых находился в горах Грейт-Смоуки, где перекрывание наблюдалось только в небольшом диапазоне высот, а другой в Бальзамических горах, где виды сосуществовали в гораздо более широкой зоне. На обоих участках обитали популяции того и другого вида и, в целом, фауна саламандр была сходной; популяции находились на одной высоте и подвергались одинаковым воздействиям. На каждом участке Хейрстон заложил семь экспериментальных площадок: на двух из них были удалены особи P. jоrdani, а двух других – особи P. glutinosus, а оставшиеся три служили контролем. Эта работа была начата в 1974 г. и в последующие 5 лет на всех площадках 6 раз в году подсчитывали число особей каждого вида; все особи были разделены на три группы: годовика, двухлетки, все остальные.
На контрольных площадках, как и следовало ожидать, из двух рассматриваемых видов гораздо более многочисленным был P. jоrdani; а на площадках, с которых он был удален наблюдалось статистически значимое возрастание численности P. glutinosus. На площадках, с которых был удален, P. glutinosus, не было отмечено соответствующего значимого увеличение численности P. jоrdani. Однако на обоих участках наблюдалось статистически значимое возрастание доли P. jоrdani среди годовиков и двухлеток. По-видимому это объяснялось возросшей плодовитостью и (или) повышенным выживанием молоди; оба эти фактора являются основными причинами, определяющими скорость размножения.
Важным моментом является то, что неблагоприятное воздействие со стороны дргугого вида исходно испытывали особи обоих видов; после удаления одного их видов, у оставшегося наблюдалось значительное увеличение численности и (или) плодовитости и (или) выживаемости. Из этого следует, что на контрольных площадках и в других местах совместного обитания эти виды обычно конкурировали друг с другом, но все же сосуществовали.
В качестве еще одного примера приведу эксперимент, выполненный одним из известнейших «отцов-основателей» экологии растений А.Г. Тенсли, который изучал конкуренцию между двумя видами подмаренника (Tansley, 1917).
Galium hercinicum – этот вид растущий в Великобритании на кислых почвах, тогда как распространение Galium pumilum ограничено более щелочными почвами. Выращивая виды по отдельности, Тенсли обнаружил, что каждый их них хорошо рос как на кислой почве из местообитания Galium hercinicum, так и на щелочной почве из местообитания Galium pumilum. Однако, при совместном выращивании на кислой почве успешно рос только Galium hercinicum, а на щелочной почве — Galium pumilum. По-видимому, эти результаты свидетельствуют о конкуренции между видами при их совместном выращивании. В конкурентной борьбе побеждает один вид, тогда как другой проигрывает настолько, что происходит его вытеснение из биотипа. Исход конкуренции зависит от условий, в которых она происходит.